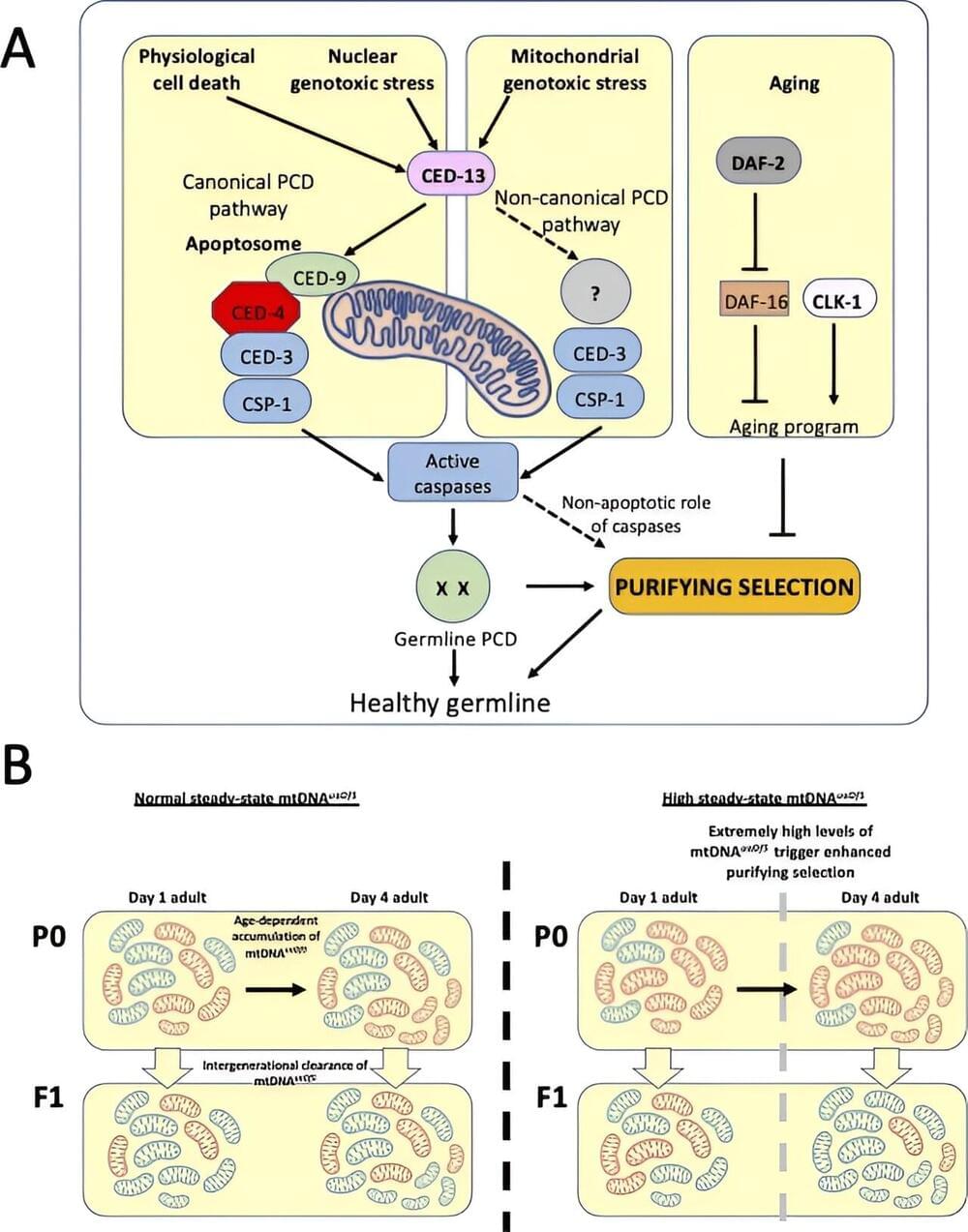
The human brain is three times bigger than a chimp’s and more spherical than a Neanderthal’s. Within a maze of bumps and grooves, neurons converse in distinct patterns that give humans unique cognitive abilities.
Scientists haven’t fully deciphered those patterns. But researchers at UT Southwestern Medical Center are determined to solve the molecular mystery of what makes us human.
In a study published in the journal Nature, they compared brain cell types and activities among humans, chimpanzees and rhesus monkeys. Human brains had more of a kind of cell that may help them adapt based on new experience and heal from injury. Certain human neurons also had more of a gene that affects language development.